Bioacoustics research
My Ph.D. in Conservation Biology focussed on two topics: kiwi calling behaviour, and conservation bioacoustics - developing tools to enhance the use of calls for kiwi conservation.
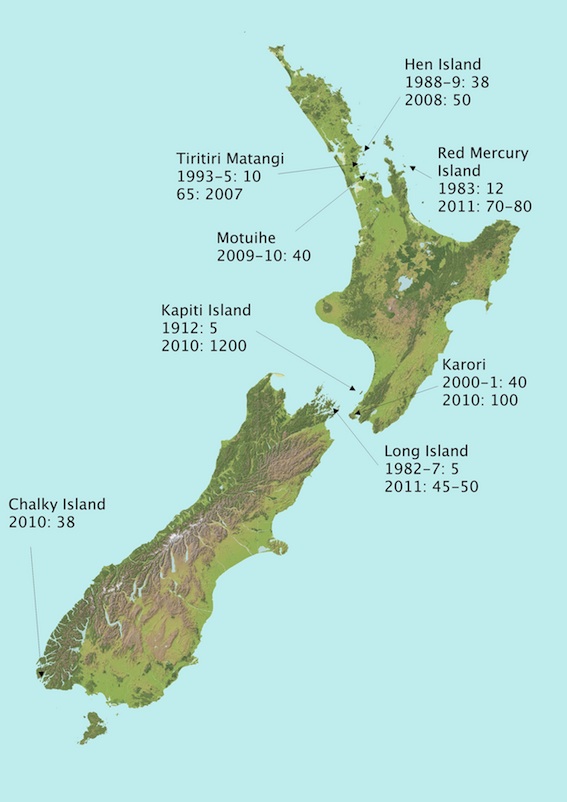
LSK populations and size, as of 2013

A female little spotted kiwi in its burrow (hover to zoom)
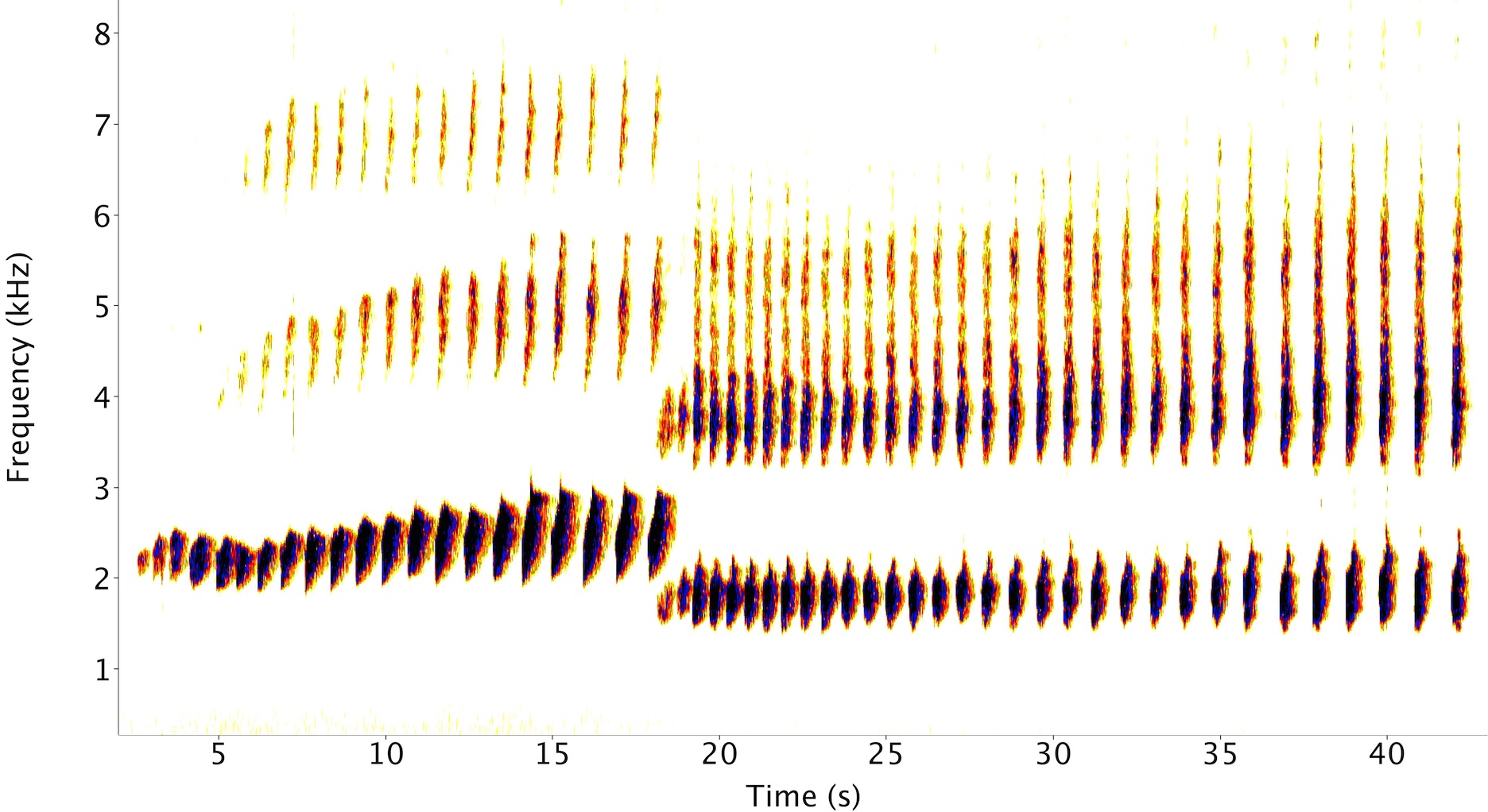
A spectrogram of a male and female little spotted kiwi call.
LSK call structure and calling behaviour
- Call description. The calls of the LSK had previously not been studied, so I first quantified the frequency and temporal properties of their calls. This is the basis for understanding more about their calling behaviour. Male LSK have higher frequency, or pitched, calls than females: this is the same for all kiwi species.
- Sexual call dimorphism. Why do males have higher pitched calls? The obvious explanation is that it’s because they’re smaller (males typically weigh about 1.1kg, whereas a female is about 1.3kg): in most animals call or voice frequency is proportional to size. Surprisingly though, in LSK there is no relation between the size of an individual and its call frequency, both within and between the sexes. See the 2013 Ibis paper below for more details.
- Call function. So if the difference in male and female LSK call frequencies isn’t a result of their size differences, what is the explanation? It’s likely that males and females have different reasons for calling. The male call travels further in the New Zealand forest, and so is better suited to longer-range functions, such as territory defence, whereas the female call, which doesn’t transmit so far, is more likely to be most commonly used for mate contact. The use of calls in duets adds further weight to these alternative functions.
- Vocal cooperation. A particularly striking feature of LSK calls is that frequency of their calls do not overlap, which cannot be explained by their size differences. Furthermore, the female call is suppressed at frequencies where it shouldn’t be - exactly where the male call is loudest. This appears to be the result of formantsamplification of certain frequencies in the vocal tract in the female call. Birds are known to coordinate their calls temporally, but this ‘vocal cooperation’ in terms of frequency has not been described in any species before. It puts strong constraints on why LSK duetwhen members of a pair call together. This feature of their calls means that both can be heard when their calls overlap, which means that they’re unlikely to be duetting for mate-guardingwhen one of the pair calls over top of the other, to make prevent it signalling so effectively to the opposite sex, but more likely to be doing so for joint territory defence, or pair bonding.
LSK acoustic identification
Do LSK have an individual call signature, like humans? The ability to identify or discriminate among individuals using just calls has strong conservation applications. It would help remove biases from call surveys when the same individuals are counted twice, and would enable remote monitoring of life history traits, such as pair-bond or territory stability, or survival.
Many other birds have individually-distinctive calls, particularly those which are unable to use sight to recognise each other - such as birds which are nocturnal, live in dense habitat, or in large colonies. Earlier studies have shown that brown kiwi (also North Island brown kiwi; Apteryx mantelli) have distinct calls, but no one has looked for this characteristic in any other kiwi species.
Further reading:
- Digby, A., Bell, B.D., Teal, P.D., 2014. Vocal individuality of little spotted kiwi (Apteryx owenii). Emu, 114 (4), 326-336
- Digby, A., 2013. Whistling in the dark: An acoustic study of little spotted kiwi. Ph.D. Thesis. Victoria University of Wellington, Wellington, New Zealand.
Non-linear phenomena
Many mammals species have structures in their calls called non-linear phenomena, which are the result of complex interactions of sound in the voice box or vocal tract. Examples of these features occur in human screams, babies’ cries, and dog barks. Their function is unclear, they tend to appear more in young, sick or subordinate animals, or in alarm calls. This suggests that they may serve to provide ‘acoustic emphasis’ to a call, to make it harder for conspecifics to ignore that individual. The use of these features may vary between species, however. Alternative hypotheses are that they aid with identification of individuals, increase vocal diversity in song birds, provide directional information, or indicate age, status or fitness of an individual.
Example of non-linear phenomena in birds are rare, most likely because few studies have looked for it. I have found that one type of non-linear phenomena, subharmonics, are common in little spotted kiwi calls. They are more common at higher frequencies, and in calls that were elicited by playbackwhen a call is played in a kiwi’s territory to simulate territorial intrusion by another kiwi. This suggests that the features provide acoustic impact, possibly to convey aggression or fitness information.
Further reading:
- Digby, A., Bell, B.D., Teal, P.D., 2014. Non-linear phenomena in little spotted kiwi calls. Bioacoustics. doi:10.1080/09524622.2013.829755.
Two-voicing
Birds have two voice boxes, or syringes, rather than the one in mammals, including humans. When birds produce sounds with both voice boxes simultaneously, this is known as two-voicing. In song birds, this may be used to add complexity to their songs. In penguin species, two-voicing has been shown to be used for individual identification. The two simultaneous sounds produce a temporal ‘beat’ that is unique to an individual. The use of this identification method varies between penguin species: Emperor and King penguins, which do not build nests, use two-voicing, but other species do not. This is because the non-nesting penguins cannot use spatial information to find their mates or young, so are more reliant on acoustic identification.
Other birds which cannot rely on visual cues to communicate frequently have two-voicing in their calls. These include birds which are nocturnal, or nest in colonies. As a bird with very poor eyesight (they have the most restricted visual fields of any bird), kiwi would might be expected to use two-voicing as a means of identifying each other. However, there is no evidence of two-voicing in little spotted kiwi calls. These kiwi may use other characteristics of their calls to identify each other, or may not even use calls for such discrimination at all.
Further reading:
- Digby, A., Bell, B.D., Teal, P.D., 2013. Non-linear phenomena in little spotted kiwi calls. Bioacoustics. doi:10.1080/09524622.2013.829755.
Vocal activity
Birds vary their calling activity with time of year, time of day or night, and weather conditions. These variation in call rates can reveal other aspects of their behaviour, such as their foraging activity, or their response to predators. For example, some nocturnal birds may call more on moonlit nights because they can find their prey more easily and so have more time for calling, whereas others will reduce their vocal output because calling makes them more easily detected by predators.
A number of studies have investigated the response of kiwi to temporal and weather conditions through the variation in call rates. However, these have all focussed on just one species, the brown kiwi, and have all been carried out using human listeners, which means that sample sizes are small. Perhaps as a result of this, the findings are mixed: for example, some studies have found that moonlight reduces call rates, whereas others have found no effect. To address these issues, I have conducted a long-term continuous listening survey for little spotted kiwi, using an automatic recorder to monitor a population all night every night for three years. I have used statistical models to determine how season, time of night, moon phase and weather conditions affect call rates in LSK. This will help further understanding of the behavioural ecology of these kiwi, but also to guide the use of call counts for kiwi monitoring (see below).
Further reading:
- Digby, A., 2013. Whistling in the dark: An acoustic study of little spotted kiwi. Ph.D. Thesis. Victoria University of Wellington, Wellington, New Zealand.
- Digby, A., Towsey, M., Bell, B.D., Teal, P.D., 2014. Temporal and environmental influences on the vocal behaviour of a nocturnal bird. J. Avian Biol., 45 (6), 591-599 . doi:10.1111/jav.00411
Duets and choruses
Kiwi pairs frequently call together in duets, either as a call and a reply, or as overlapping calls. Also, members of the same sex will often call and answer each other in a sequence, or chorus. It is unclear why kiwi call like this, but it’s probable they do so for the similar reasons as other birds. The main theories for duets in birds (and for other animals) are:
- territory defence: when a pair join together to defend their territory - “two voices are better than one”
- pair contact: so the members of a pair can find each other
- pair-bond reinforcement: calling together can reinforce the bond between a male and female, and perhaps assist in synchronising reproduction.
- mate-guarding: a member of a pair can call at the same time as its mate to prevent it attracting the attention of other potential mates.
Choruses in kiwi often involve males calling and answering each other, which may mean that they are marking their territories, or perhaps engaging in intra-sexual competition.

A little spotted kiwi chick, about a week old
Chick calls
I have recorded LSK chicks in their nests, from hatching through to several weeks of age (they typically leave the nest after about 4 weeks, but can stay in the nest for up to 3 months). The chicks are surprisingly vocal, particularly when the male returns to the nest (in LSK and brown kiwi, only the male incubates the egg and shares the nest burrow with the chick). But what is the purpose of these vocalisations? To help determine this I am investigating the structure of the chick calls, and how they change with the presence of the male, and as the chick grows older. Initial results show that LSK chick calls are rich in complex vocal features absent from adult calls.
Further reading:
- Digby, A., 2013. Whistling in the dark: An acoustic study of little spotted kiwi. Ph.D. Thesis. Victoria University of Wellington, Wellington, New Zealand.
Conservation bioacoustics
All kiwi species are threatened with extinction (LSK are officially listed as near-threatened because their population is increasing as they are all in predator-free habitats, but there are concerns about their low genetic diversity). Since kiwi are cryptic, nocturnal and can live in dense forest habitats, they are hard to monitor. Fortunately they are vocal: in all five species both males and females emit loud whistle calls that can be heard for hundreds of metres. So call surveys are an important method of monitoring of kiwi populations. These “call counts”, many conducted by volunteers, are carried out throughout New Zealand to help kiwi conservation. They are used to map kiwi distributions, detect remnant populations, and assess the effectiveness of conservation management methods such as predator control and advocacy.
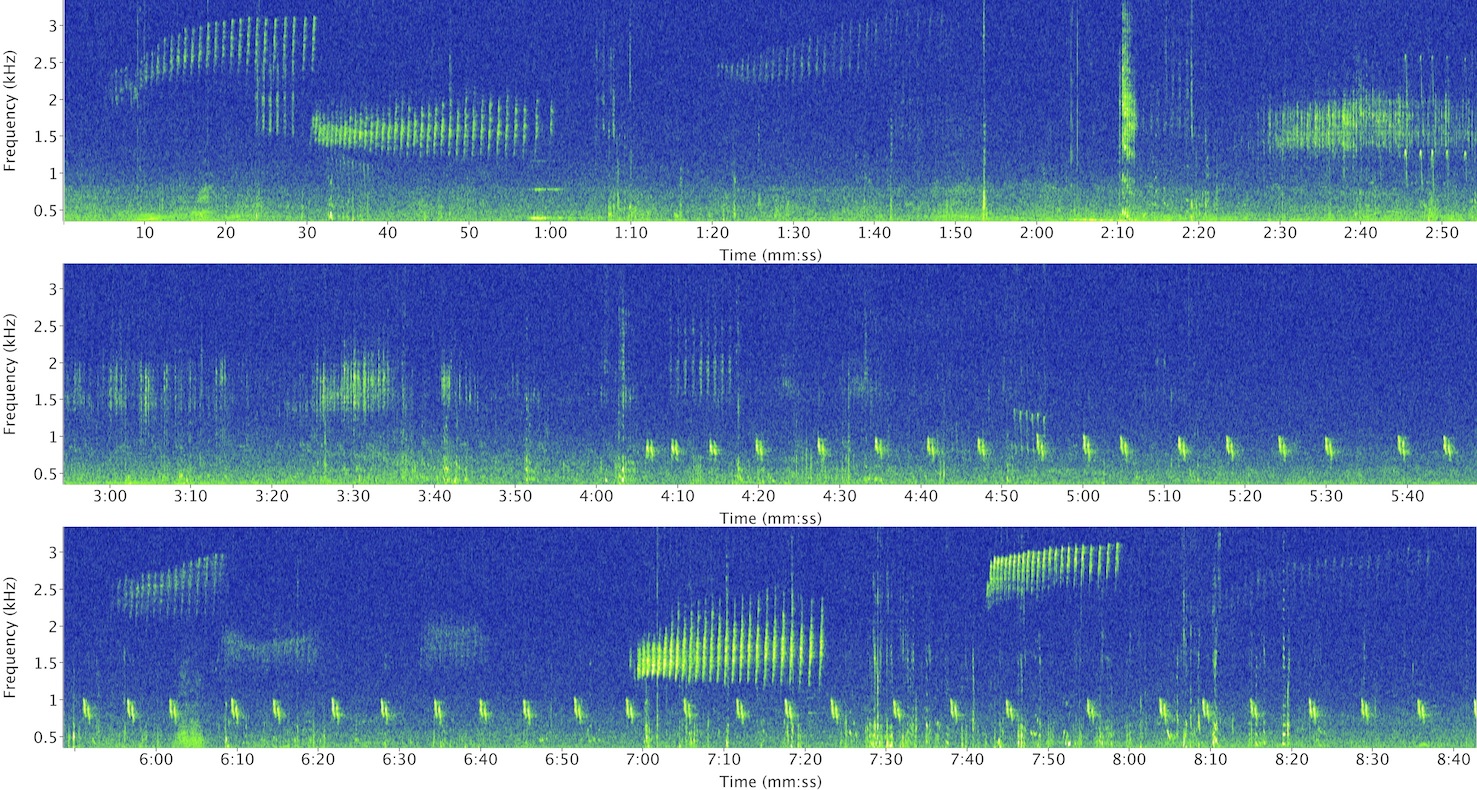
A spectrogram from an autonomous recorder
Autonomous monitoring
Although call counts using observers “in the field” are a key part of kiwi conservation, they are prone to several problems. First, hearing ability differs between people, and each person’s ability to detect kiwi will vary with time - either from year to year, or even throughout a single count. With no way of checking what they just heard, observers can misclassify calls, commonly mistaking ruru (a native owl) or weka (a rail) for kiwi. These counts are also usually conducted over short time frames (1-2 hours on just a few night), and usually at the beginning of the night at the same time of year. They are also carried out under fine weather conditions, and usually only in accessible habitats. Call counts using field observers are therefore subject to a number of biases, and kiwi calling behaviour is largely unknown outside of the times, weather conditions and places in which the counts take place.
Using automatic recorders can resolve many of these problems. They are widely available, relatively inexpensive, and can be scheduled to record each night for long periods. However, recorders are usually less sensitive than a human listener, so cannot sample such a large area. I have estimated that a typical model covers about 80% of the area a person can hear. To extend the monitoring area, more recorders can be used. The recorders normally do not provide the spatial information (where the kiwi are calling from) that a person in situ can provide. They therefore offer an alternative to field observers, but are perhaps best used in conjunction with “manual” counts.
The recordings are usually processed using a spectrogram - a visual representation of the sound. A person familiar with spectrograms can conduct kiwi counts quite quickly, processing the data at a rate of around 2-5 minutes per hour of recording. Doing counts in this way is therefore much more efficient, and accurate, albeit less sensitive. The recordings are a permanent record of the count, so can be reanalysed if required.
A further possibility is to process the spectrograms automatically, using software to detect and classify each sound. This field is still in its infancy in terrestrial ecology, and automatic detectors further decrease the recorder sensitivity. However, as these algorithms improve, the efficiency of acoustic monitoring will dramatically increase. Myself and co-authors compared LSK call counts using field observers, spectrograms and fully-automatic classification, and found that the fully-automated method covered about 40% of a manual survey, and was over 30 times more efficient.
Further reading:
- Digby, A., Towsey, M., Bell, B.D., Teal, P.D., 2013. A practical comparison of manual and autonomous methods for acoustic monitoring. Methods in Ecology and Evolution 4, 675–683.
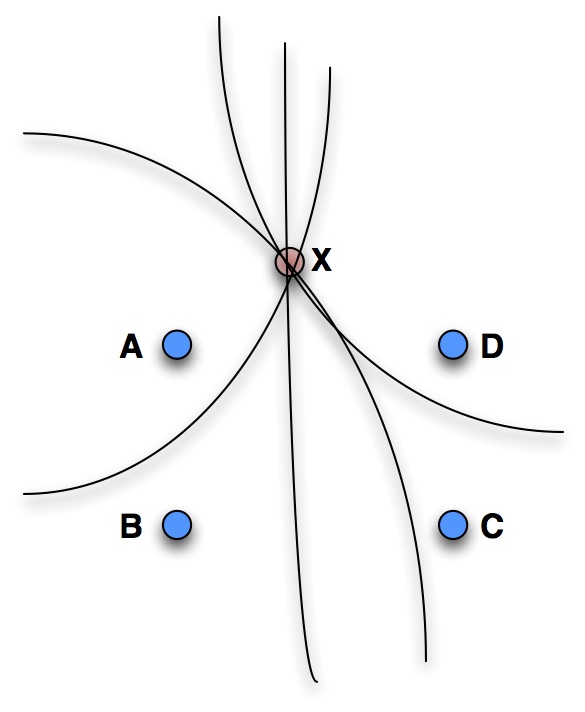
Multilateration: the time delay between each pair of microphones (A-D) describes a hyperbola. If multiple pairs are combined these intersect at the source location (X).
Microphone arrays
Using multiple microphones in an array allows the location of calls to be determined, as well as recording the calls themselves. One way of achieving this is through multilateration. The time delay between a sound reaching each microphone in a pair determines that the sound source lies on a hyperbola (curve). By combining multiple pairs, the hyperbolae intersect at a point, providing an estimate of the call location. With four or more microphones, the source position can be estimated in three dimensions.
Microphone arrays have been used for many years in marine mammal monitoring, but until recently have been relatively under-exploited in terrestrial ecology. Experiments have shown that arrays can determine the position of calling birds to sub-metre accuracy, even in thick forest habitats.
Most of these experiments have used wired arrays, which require hundreds of metres of expensive cables which are difficult to deploy in the bush. Also, nearly all have used custom-built equipment. No one has also used microphone arrays for kiwi, despite their loud, repeated and frequency modulatedthe frequency of the notes, or syllables, changes rapidly with time calls being ideally suited for localisation methods. For my Ph.D. I developed and tested microphone array methods for kiwi monitoring, using commercially-available wireless recorders.
- Digby, A., 2013. Whistling in the dark: An acoustic study of little spotted kiwi. Ph.D. Thesis. Victoria University of Wellington, Wellington, New Zealand.